Home
Modeling
human expertise on a cheese ripening industrial process
Cheese
Ripening
In
order to make so-called "model cheeses", experimental
procedures in laboratories use pasteurized milk inoculated with
Kluyveromyces
marxianus
(Km), Geotrichum
candidum
(Gc), Penicillium
camemberti
(Pc) and Brevibacterium
auriantiacum
(Ba) under aseptic conditions.
During
ripening, these soft-mould cheese behave like an ecosystem (a
bio-reactor) extremely complex to be modeled as a whole, and where
human experts operators have a decisive role. Relationships between
microbiological and physicochemical changes depend on environmental
conditions (e.g. temperature, relative humidity ...) and influence
the quality of ripened cheeses. A ripening expert is able to estimate
the current state of some of the complex reactions at a macroscopic
level through its perceptions (sight, touch, smell and taste).
Control decisions are then generally based on these subjective but
robust expert measurements. An important information for parameter
regulation is the subjective estimation of the current state of the
ripening process, discretised in four phases:
|
Phase
1
is characterized by the surface humidity evolution of cheese
(drying process). At the beginning, the surface of cheese is very
wet and evolves until it presents a rather dry aspect. The cheese
is white with an odor of fresh cheese.
|

|
|
Phase
2
begins with the apparition of a P.
camemberti
coat (i.e the white-coat at the surface of cheese), it is
characterized by a first change of color and a "mushroom"
odor development.
|

|
|
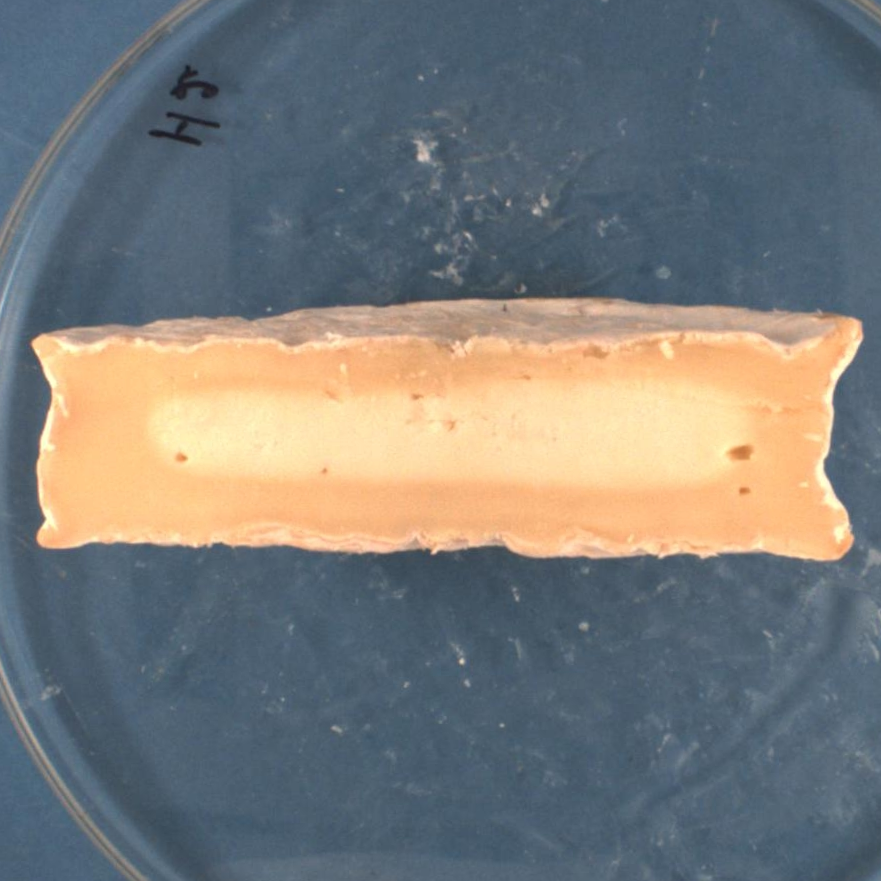
Phase
3
is characterized by the thickening of the creamy under-rind. P.
camemberti
cover all the surface of cheeses and the color is light brown.
|
|
|
Phase
4
is defined by strong ammonia odor perception and the dark brown
aspect of the rind of cheese.
|

|
These
four steps are representative of the main evolution of the cheese
during ripening. The expert's knowledge is obviously not limited to
these four phases, but they help to evaluate the whole dynamics of
ripening and to detect drift from the standard evolution.
Evolutionary
Algorithms: Cooperative co-evolution techniques
|

|
Cooperative
coevolution
strategies
rely on a formulation of the problem to be solved as a cooperative
task, where individuals collaborate or compete in order to build a
solution. They mimic the ability of natural populations to build
solutions via a collective process.
Instead
of dealing with a coevolution process that happens between
different separated populations, we use a different implementation
of cooperative coevolution principles, called "Parisian
approach", that uses cooperation mechanisms within a single
population. It is based on a two-level representation of an
optimization problem, in the sense that an individual of a
Parisian population represents only a part of the solution to the
problem. An aggregation of multiple individuals must be built in
order to obtain a solution to the problem. In this way, the
co-evolution of the whole population (or a major part of it) is
favoured instead of the emergence of a single best individual, as
in classical evolutionary schemes.
|
Genetic
Programming
|
Genetic
Programming (GP) is a specialization of genetic algorithms where
each individual is a function, represented as a tree structure.
Nodes
Every
tree node has an operator function: +,-,/,*, ...
Every
terminal node has an operand: a constant or a variable
It
is then easy to apply genetic operators to these trees in an
evolutionary algorithm.
|
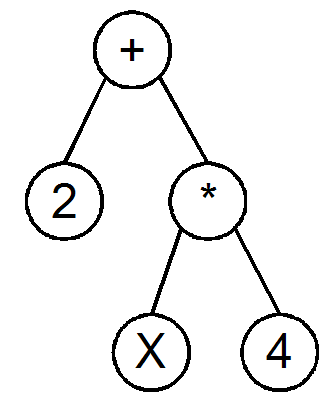
|
GP
is used here to search for a convenient formulas that estimate the
phase at time t,
knowing micro-organisms proportions at the same time t,
but without a priori knowledge of the phase at the time t-1.
In a first classical GP approach, the phase estimator is searched as
a single best "monolithic" function and then we use a
cooperative scheme to split the phase estimation into four combined
(and simpler) "phase detectors".
Phase
estimation using a classical GP
The
derivatives of four variables will be considered, namely the
derivative of pH
(acidity), la
(lactose proportion), Km
and Ba
(lactic acid bacteria proportions), for the estimation of the phase.
The GP will search for a phase estimator Phase(t),
i.e. a function defined as follows:
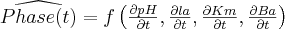
The
fitness function, to
be minimised,
is made of a factor that measures the quality of the fitting on the
learning set, plus a "parsimony" penalisation factor in
order to minimize the size of the evolved structures in order to
avoid bloat. It favours evolution of structures with 30 to 40 nodes.
Moreover, it is divided by the number of variables involved in the
evaluated tree in order to favour structures that embed all four
variables of the problem.
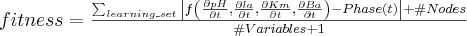
Phase
estimation using a Parisian GP
Instead
of searching for a phase estimator as a single monolithic function,
phase estimation can actually be split into four combined (and
simpler) phase detection trees. The structures searched are binary
output functions (or binarised functions) that characterize one of
the four phases. The population is then split into four classes such
that individuals of class k
are good at characterizing phase k.
Finally, a global solution is made of at least one individual of each
class, in order to be able to classify the sample into one of the
four previous phases via a voting scheme.
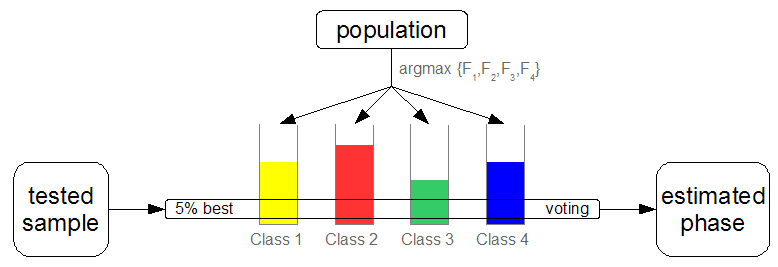
We
now search for formulas I()
with real outputs mapped to binary outputs, via a sign filtering:
(I()>0)
-> 1
and (I()<0)
-> 0.
The functions (except logical ones) and terminal sets, as well as the
genetic operators, are the same as in the previous global approach.
Using the available samples of the learning set, four values can be
computed, in order to measure the capability of an individual I
to characterize each phase (i.e. if I
is good for representing phase k,
then Fk(I)
is positive for k
and negative for the other values.):
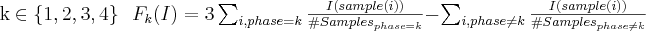
The
local adjusted fitness value, to
be maximised,
is a combination of three factors:
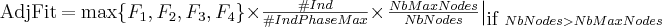
The
first factor is aimed at characterising if individual I
is able to distinguish one of the four phases, the second factor
tends to balance the individuals between the four phases
(#IndPhaseMax
is the number of individuals representing the phase corresponding to
the argmax
of the first factor and #Ind
is the total number of different individuals in the population) and
the third factor is a parsimony factor in order to avoid large
structures.
At
each generation, the population is shared in four classes
corresponding to the phase each individual characterises the best.
The 5%
best of each class are used via a voting scheme to decide the phase
of each tested sample. The global fitness measures the proportion of
correctly classified samples on the learning set and is distributed
as a multiplicative bonus on the individuals who participated in the
vote
Experimental
analysis
Available
data have been collected from 16 experiments during 40 days each,
yielding 575 valid measurements. It is to be noticed that collecting
these experimental data was a long and difficult process, so these
resulting data sets are often uncertain or even erroneous. For
example, a complete cheese ripening process last 40 days, and some
tests are destructive, i.e a sample cheese is consumed in the
analysis. Other measurements require to grow bacterias in Petri
dishes and then to count the number of colonies, which takes a lot of
time.
Experiments
show that both GP outperform other available methods (multilinear
regression and Bayesian network) in terms of recognition rates.
Additionally the analysis of a typical Parisian GP run shows that
much simpler structures are evolved: The average size of evolved
structures is around 30 nodes for the classical GP approach and
between 10 and 15 for the Parisian GP.
It
has also to be noted that co-evolution is balanced between the four
phases, even if the third phase is the most difficult to characterize
(this is in accordance with human experts' judgement, for which this
phase is also the most ambiguous to characterize)
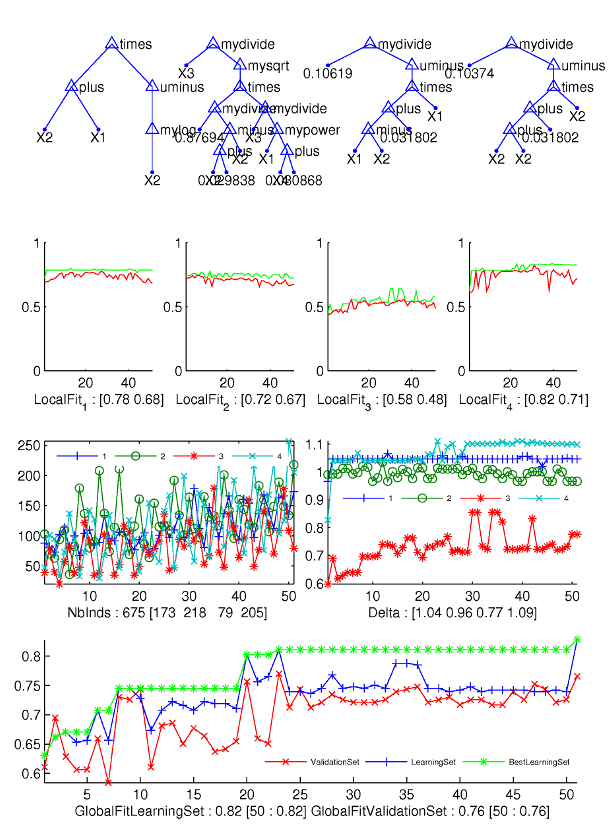
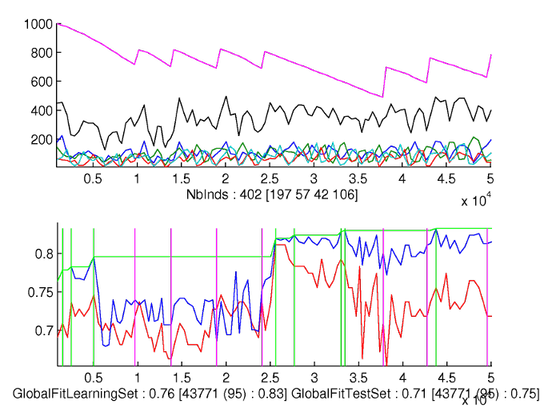
A
typical run of the Parisian GP:
First
line: best evolved trees of each class
Second
line: the evolution with respect to generation number of the 5% best
individuals for each phase: the upper curve of each of the four
graphs is for the best individual, the lower curve is for the "worst
of 5% best" individuals.
Third
line left: the distribution of individuals for each phase: the
curves are very irregular but numbers of representatives of each
phases are balanced.
Third
line right: discrimination indicator, which shows that the third
phase is the most difficult to characterize.
Fourth
line: evolution of the recognition rates of learning and test set.
The best-so-far recognition rate on learning set is tagged with a
star.
Variable
population size strategies in a Parisian GP
|
Despite
of local elitism and bonus mechanisms, the global fitness is not a
monotonically increasing function. In particular, it often happens
that a generation notably improves the global fitness, while the
generations that follow are not able to keep it. To avoid this
undesirable effect, a variable sized population Parisian GP
strategy is experimented, using adaptive deflating and inflating
schemes for the population size.
The
idea is to group individuals with the same characteristics into
"clusters" and remove the most useless ones at the end
of every generation while periodically adding "fresh blood"
to the population (i.e. new random individuals) if a stagnation
criterion is fulfilled.
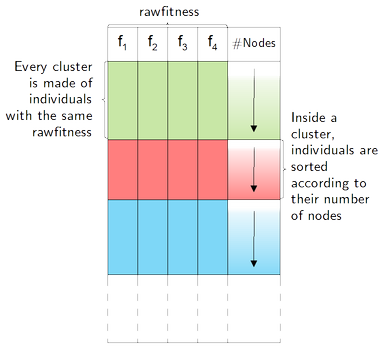
Individuals
having the same rawfitness
are grouped into clusters. Then, inside each cluster, individuals
are sorted according to their number of nodes. The first and best
one is the one with the smallest number of nodes.
Useless
individuals elimination allows to decrease the population size: An
individual is considered as useless if it belongs to a big cluster
and has a large number of nodes. The elimination rule depends on
two parameters (tokeep
and toremove):
if a cluster has less than tokeep
individuals, they are all kept, and if it has more, only the last
toremove,
having the largest number of nodes, are removed.
|
|
|
|
In
order to avoid stagnation due to over-specialisation of the best
individuals, we propose to periodically add "fresh blood"
to the population (i.e. new random individuals) if a stagnation
criterion is fulfilled. The corresponding algorithm uses one
parameter denoted toinsert,
typically set to a lower value than tokeep
In
this way, if a cluster of the old population is empty or has not
enough elements according to a stricter rule than during the
elimination process, it gets new elements.
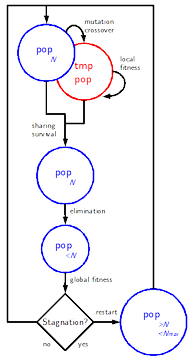
The
full inflating-deflating scheme is made of the following steps:
mutations
and crossover
yield a temporary population tmppop
local
fitness
is computed on the temporary population: localfitness(tmppop)
adjusted
fitness
is computed via sharing: sharing(pop+tmppop)
selection
of the N
best individuals: pop=survival(pop+tmppop)
elimination
of the useless individuals: pop=elimination(pop)
global
fitness
computation of the global fitness of the population:
globalfitness(pop)
partial
restart
if a stagnation criterion is met: pop=restart(pop)
|
With
the deflating+inflating scheme, there are improvements of the global
fitness all along the generations. The final recognition rate on the
learning set is better than with the two other schemes. As far as the
size of the population is concerned, one can observe the cycles of
deflations and partial restart. The population is still quite
balanced between the four classes, and the number of unique
individuals is also quite stable.
This
first attempt to manage varying population sizes within a Parisian GP
scheme show the effectiveness of the population deflation-inflation
scheme in terms of computational gain and quality of results on a
real problem. The deflating scheme allows to obtain the same result
as the fixed-size population strategy, but using less fitness
evaluations. The deflating-inflating strategy improves the quality of
results for the same number of fitness evaluations as the fixed-size
strategy.
More
generally, the deflation-inflation scheme has two major
characteristics: a clusterisation-based redundancy pruning and a
selective inflation, which tries to maintain limited-size clusters
with low complexity individuals. These two concurrent mechanisms
tends to better maintain low complexity individuals as well as
genetic diversity. These characteristics may actually be transposed
to classical GP or EAs, in particular to limit GP-bloat effects.
Home